Reciprocal Causationʹ, the Biosphere, and Intelligent Design
RECIPROCAL CAUSATIONʹ, THE BIOSPHERE,
AND INTELLIGENT DESIGN
Virgil Warren, PhD
Introduction
The following observations evaluate general evolution in the organic realm with particular attention to the animal kingdom. They stress features of living reality based on reciprocal causationʹ, its necessary accompanying features, and its implications in contrast to all forms of straight-line causal series. The following diagrams picture various causal formats.
 Types
of reciprocal causation
Types
of reciprocal causation
 Types
of straight-line causal series for contrast purposes
Types
of straight-line causal series for contrast purposes
Comments related to these causal formats appear in the explanations below.
PART I
Question #1
Can material nature unaided put in sequence the origin of reciprocatingʹ parts that each together produce all the effects that are necessary for the ongoing viability of the whole organism and its other parts? [system level per kind of entity]
Essential feature in epitome: heart-lung-heart reciprocation
the heart pumps necessary nutrients to . . . and the lungs,
which provide the oxygen necessary for . . . and the heart.
Embedded implications somewhat overlapping, redundant, and variously noted
The given: LIVING BODIES comprised of real living, acting, reciprocallyʹ
interdependent, complementary parts A kind of living perpetual motion machine
Origination
Boundary limit: reciprocal causationʹ within an outer boundary
This essay has added ʹ (the “prime” mark) to “reciprocal” and “reciprocal causation” to specify a reciprocal causation that receives back other than what it produces—that is to say, something other than increased degree of the same thing as in feedback between an amplifier and guitar or the echo effect in its variant applications.
Completeness limit: reciprocal causationʹ within a bottom limit (irreducible)
No needed parts missing from this entity/system
The level of the whole transcends non-living prebiotics.
The whole transcends chemicals and chemical clusters that would supposedly “come to life” when stimulated in some way, especially if conceived of as combining gradually.
The entity has parts that are individually alive within it, parts that, if separated from the whole, would lose viability (die and rot).
Each part must be alive/functioning, not just chemically present.
Viability limit: the particular kind of requirement within the limit
Reciprocal causationʹ applies to life in its origination and continuation.
Viability applies to the parts and the whole that the parts comprise.
Viability limit applies to the degree of variation within parts and their relationships as bounded by constraints of functionality and reproduction—color, size/degree, shape, proportionality, along with presence of non-essential elements (hair, nails).
The process has given, observed knowns.
No postulated splices
No specter of free-floating, functional parts able to come together randomly to form a functioning whole—as might be conceivable in inorganic chemistry; the parts would not be living and so would be non-functional aside from participation in the living entity.
Parts are unlike, say, individual chemicals or 2-by-4s, able to exist separately “on their own” while potential for combining with others to a desired effect. Neither the parts nor any combination of them would be functional. The parts would not be functional because they would not yet be receiving from the other parts of the “whole” what was necessary for them to live and function; the “whole” would not be living because it did would not yet be receiving from the still uncombined part what that part would need to be contributing to make the whole viable and functional. So, all parts would have to be present for functionality purposes, and parts not already present would be in a state of deterioration and decay relative to their necessary condition for viability purposes (doubly impossible).
All parts operational from the first; no “becoming” of parts or the whole
All parts connected harmoniously from the beginning (coherence)
No disconnects, blockage, gaps, misdirects
All parts able to “nest” with the rest
All parts originating simultaneously (virtually) as per the viability limit both higher and lower
Simultaneous origin of reciprocatingʹ parts
Gradual origin of each part eliminated
Serial origin of parts eliminated
Brief time for originating the whole entity and its necessary parts
Viability limited to minutes without oxygen (lungs) and to days without nutrients (heart)
Urgent origination in contrast to gradual origination or accumulation of parts
The inherent impossibility in the real world of sequencing the origin of functioning necessary parts reciprocatingʹ in a closed loop of living parts—reciprocal causationʹ in a closed living loop
Viability limit requires immediacy, sufficiency, and consistency of what each previous organ in the loop supplies.
Proximal time and place of origin
All parts of each entity originating at the same time and place
No alternative sources—real or postulated, internal or external—for what each necessary part produces (boundary limit as to inclusion and exclusion)1
Specificity of each part’s effect within the body (boundary limit)
No part functionally “first”
Every part receiving from all the others what it needs in ongoing fashion for living and supplying what is necessary for them in ongoing fashion
Every part capable from the beginning for making its contribution before receiving from the others (capacity), but unable to keep producing without receiving the others’ ongoing contributions (operation)
Provision for parts beyond minimum functionality: two ears, two eyes
Provision for parts beyond survival necessity
Double provisions for back-up: kidneys, lungs, testes
No apparently irrelevant, useless parts
Contra imaginary vestigial or nascent organs
Extreme complexity of parts and their combinations within the whole
Which dictates against random, gradual, serial origin of parts
Which dictates against undesigned systems and operations
Perpetuation
All parts functioning sequentially
All parts functioning simultaneously
Every part operating in the context of entropy (death, viability limit)
Different kinds of parts contributing different kinds of results, all of which results are
necessary for the viability of the whole and each other
Parts variously functioning involuntarily, semi-voluntarily, or voluntarily
All the effects of all the other necessary parts come back around to enable “this one”
(serial reciprocationʹ)
All products of each part being of a kind that can meet the needs of the other parts to
exist and/or to produce what they produce
All parts producing sufficiently (amount/degree, rate, immediacy, consistency) what
all the other parts need from them for living (viability) and to producing their
necessary effects (functionality)
Sufficiency that includes variability in functional requirements
Each part producing something other than itself: reciprocal causationʹ
Not as in “shedding” or rubbing off
Each part able to receive what the preceding parts supply
Each part receiving sufficiently everything it needs in amount, rate, immediacy, and
consistency
Each part receiving what is other-in-kind from its own products
All parts necessary: “irreducible minimums”
Reciprocalityʹ shows why the most to least complex set of minimums is irreducible within its whole.
Nothing detrimental, deleterious, interrupting, or lethal in the operational context of the system or its parts
Ongoing process, active and progressive
Not coming to a soon “dead end,” stopping, or halting prior to reproductive operation of the whole
Continuing to have ongoing, productive functionality
A needs B in order to keep being A; B needs A in order to keep being B; likewise with C, F, G, T, etc.
Accompanying instincts and behaviors for feeding, protection, self-preservation, and reproduction
The reciprocatingʹ parts in hierarchy complexities
The whole sits atop hierarchical levels of complexity arranged downward toward the basic parts (quarks in protons-and-electrons, in molecules, in chemicals, in sets within cells that form functioning units arranged upward in levels toward the whole)
Degrees of healing/self-correcting among the parts, kind of add-on to necessity
The complete set of reciprocatingʹ parts also incorporate many straight-line processes.
Accounting for all bodily systems and what enables them:
Sensory, nervous, musculature, ambulatory, skeletal, endocrine, digestive, circulatory, in-take, elimination, reproductive—each of which must already be coherent within each variant entity both between the parts and between the subunits that in turn make up the aggregate, hierarchical arrangement all the way down to the cellular level and below
High-level complexity comprised of reciprocationʹ in addition to various types of straight-line causality that may also be present
The reciprocal causationʹ pattern, epitomized under heart-lung reciprocalityʹ, gets multiplied endlessly when all the bodily systems are seen together. Reciprocal causationʹ removes statistics (probabilities) from consideration by noting impossibility by the nature of the case in the natural world.
In addition to necessary parts of the whole, there are parts that in themselves are too complex to originate gradually by mutation and natural selection. Note also the irreducibly complex set at the cellular level, hence, also of intelligent design.2
Involving more than straight-line causal series (See diagrams in the introduction.)
Domino effect direct and indirect
Echo effect
Something that increases the number or degree of what first went out
Like feedback between a guitar and amplifier
Like the warming feedback loops in climate-change theory
Like love in response to love
Static interdependence
As in attached row houses
Dead-end formats,
Straight-line or combinational, even where what combines produces a different
result from the components
Hegelianism
As a kind of open-ended, mutually adjusting combinationalism
Gestalt
A whole that is more than the sum of its parts; completing, dead-end
Symbiosis between two systems already functional
Parasitic: one benefits, the other is harmed (The host dies from the parasite.)
Commensal: one benefits, the other is not affected (Both can exist separately.)
Mutual: both benefit (bees and plants that are pollinated only by bees)
Concluding comments: living organisms cannot originate from any form of straight-line causation, nor can they continue functioning that way, at least in the animal kingdom. Among all living organisms, “kinds” must have originated as fully functional, internally reciprocatingʹ sets.3
So, from what we can tell or postulate scientifically, the answer to Question #1 is “No.”
Question #2
Can nature then produce complementing variants of Question-#1 entities that reciprocallyʹ interact to replicate both variants intergenerationally? [individuals within kind of entity]
Embedded realities and implications
The given: TWO SEXES, variant and biologically complementary for reproduction
through unification
Diverse bodies in a reciprocatingʹ, combinational set
(cp. in a different connection “woman from man and man by woman”
1 Corinthians 11:11-12)
Deals with organisms above the fission level of reproduction
Applies to the fusion reproduction level
Proximal time and place of origin
Same point of origination in time and place for both variants
No alternate source for the next generation in a bisexual system than the union of
interbreedable pairs and the dual-variance result of that union (male-female)
Operational only between the paired variants of each kind
Bounded bisexual interbreeding units amidst an astronomical number of other such kinds
Coordinated drives and receptivity
Accompanying instincts and behaviors for reproduction and “parenting”
Nothing detrimental or lethal in the context of operation between the paired entities
Ongoing intergenerational perpetuation as dependent on environmental considerations
Every generation operating in the context of short-term entropy (death)
Concluding comment: The individuals in all living entities must come from “parents” that have the abilities, instincts, and behaviors to provide everything necessary for the offsprings to develop into independent, living, replicating entities (DNA). The “parents” can provide for the temporary, various circumstances in which those different entities can do so (womb, pouch, egg).
So, the answer to Question #2 is also “No.”
The impossibilities implicit in Question #1 plus #2 combine to highlight many hierarchical levels of impossibility for explaining the biosphere by aggregates of causes within physical wholes. With reciprocal causation’sʹ eliminating irreducible minimums at the level of the whole in living entities (on top of internal compenetrating systems and irreducible complexities within biochemical building blocks), the origins issue changes from statistically improbable to inherently impossible as figured relative to the sure characteristics of physical nature. Reciprocal causationʹ eliminates the probability option.
Highlighting reciprocal causation (also called a “loop”) at the level of the whole is probably easier to see than other irreducible complexities at the cellular and molecular levels of living entities. Impossible in these matters contrasts with improbable as further discussed in PARTS II and III.
PART II
Not being able to appeal to alternate sources includes discounting biological evolution from a previous kind in a postulated tree of life. PART I focuses on the parts of a living whole and observes that there is no alternate surrounding source for any necessary part or its product. Biological evolution, however, proposes to get that “external” origin from a previous source through the parenting portal. That move endeavors to replace category with continuum, kind with degree. While descent opens up a natural source, it does so with a larger downside than the original problem of getting a new kind of entity: it introduces a previous whole, one that already has all its parts (category), not just maybe one or two that the new entity may need or may need upgraded. That scenario would mean numerous, incremental, serial/sequential, coordinated, random, positive, mutations-plus-natural-selection on all the parts that make up the previous entity in order to adjust it en masse to the new entity.
(1) That process would need to happen while keeping the entity functional in all its parts in all its stages of progress. That would be like modifying a model T into a Cadillac while it was driving down the road. Viability limit plus reciprocal causationʹ plays large here because, unlike an inorganic development (like an automobile redesign), the earlier functioning entity cannot comparably lie dormant indefinitely in a non-living, intermediate form until something randomly advances it forward another step toward some unforeseen viability point—and then resurrect to proceed again: “Dead men don’t rise.” The previous entity dares not lose its viability, and the subsequent entity cannot do without it—obviously.
(2) The modifications would have to take place in coordination across all the innumerable necessary parts in order to bring into a new life form.
(3) Each mutation stage on each of the modifying parts would have to be beneficial to be selected for, hence, not neutral or deleterious like most mutations are.
(4) Each part’s innumerable mutations would have to progress in series while, in coordination with all the other parts’ innumerable mutations, progressing in series toward a common goal within each part and for the sum of parts that cohere in the continuously functioning set.
(5) Even more stymieing, how would evolution en masse work with a mosaic like the famous platypus, which combines features from across supposed evolutionary lines? The platypus is a semiaquatic mammal that lays eggs and has webbed feet and a kind of bill like a duck, has fur like an otter and a broad flat tail like a beaver. A designer could decide to mix templates once in a while—out of sheer delight in doing so, but physically evolving such a mixture of disparate features randomly from parallel streams of descent defies imagination, let alone experimental demonstration. Mosaics fit better a web of life forms than a tree of life.
(a) The argument calls for a negative burden of proof for denying general evolution (“disproof”), but proper reasoning calls for the claimant to shoulder the positive burden of proof.
The closest evidence Darwinian theory appeals to is structural similarity between templates of life forms (body plans), which in turn are loosely correlated with fossil remains in geological stratigraphy. We say “loosely” because the whole geological column of levels in superposition does not appear in any one place (gaps), and because disconformities from standardized sequence also appear. Leaping over gaps in the structure and disconformities in the format, that standardized sequence originates largely as a mental simple-to-complex construct projected onto fossil remains, which relies again on mutation-selection. The specific connections and correlations are postulations about the past, not empirically observable in the present. So, the geological argument and the structural similarity argument both come from the same metaphysical assumption: mutation-selection arranged in a simple-to-complex process. The structural-similarity observation and geological-stratigraphy interpretation are taken to correlate.4
(b) The argument appeals to imagination (metaphysical) rather than observations about reality, where the evolution would be occurring (physical). The connections between homologous components are pictured as lying within the realm of the entities rather than outside their realm in the mind of a designer.
The argument offers postulations about the invisible past, about processes too slow to watch, about changes too small to observe, especially at the cellular and molecular levels. It commits a “category error”: metaphysical for physical. The theory is unobservable, invisible, and without prediction power; hence, it is not testable and therefore not scientific. In other words, its apparent strength comes from reasoning in the form,
“You can’t disprove what I can imagine;
so believe it happened.”
But Darwin’s theory itself offers no positive evidence (a) for random mutations lining up in series toward goals, or (b) for accumulating elements into complex sets functional only after becoming complex sets, or (c) for spontaneously generating a set of reciprocal causesʹ in a closed living cycle. The whole point behind proposing randomness is to replace mind-intentionality-authority with unguided operation of universal, mindless physical “laws” so as to mimic design with chance. Mutations, however, are most likely neutral or negative. Even when positive, they are positive in the short term but delimiting (degradative) in the long term because they destroy a possibility by removing, changing, or rearranging a DNA structure away from the original “in the wild” (Michael Behe, Darwin Devolves: The New Science About DNA That Challenges Evolution, pp. 225-552). The original form of the entity cannot be regained.
Besides, previous and present entities do not necessarily have the same number of necessary parts with the same size, shape, sequence, and arrangement; or cohere in the same way. Natural selection would work against nascent organs, e.g., as non-functional extras like tumors; so, mutation-selection would not lead to new organs that a new entity that we know exists would need to obtain (from one gestalt to a different gestalt). Mutation-selection would work against vestigial organs by eliminating something necessary for viability in the previous entity to descend from (from non-gestalt to gestalt). A “gestalt” is a whole, which is more than the sum of its parts, and the main factor beyond “the sum of parts” is unity (coherence) unto functionality. From what we do know, the biosphere as an irreducibly complex functional system requires an intelligent designer that works “from scratch” or at least from the nearest previous or other entity according to an array of fundamental templates. The designer arranges parts to accomplish a purpose.
In reality, research shows that the possible variations within a kind are limited to the built-in potential variants of each entity’s DNA (internal). Truly new functional forms do not arise from unintelligent environmental causes (external).
To complete the set from whole entities (PART I) through organs (PARTI II), attention needs to shift on down to the molecular level because that is where the building blocks of life lie and mutations occur, so that is the level Darwinism banks on being able to account for biological origins and perpetuations (Michaelk Behe, A Mousetrap for Darwin, p. 320).
This level not only requires modification en masse for all the countless necessary organs/parts, it requires modification en masse of the hundreds of thousands of amino acids, nucleotide bases, proteins, genes, chromosomes, DNA base sequences, etc., in the cells. That must occur, again, in a way that does not lead to dead ends for those millions of sub-cellular items and the cells that encompass them while keeping the organs individually functional and the whole entity viable as they somehow get to entities that are incompatible with them reproductively.
However imaginatively plausible the general Darwinian theory may seem, it diminishes inexorably the more unlike the templates (design patterns) become. So to speak, going from a wren to an albatross by small, unguided steps is hard enough to imagine, but imagining a path of connection between a snake and a whale is something else. It is hard enough to imagine; it is impossible to demonstrate.
PART III
If causes internal to the natural order cannot explain everything in the natural order, then the natural order itself requires causation external to itself to account for itself (1) to get started as well as (2) to be supplemented along the way thereafter. We cannot observe that external cause itself, but we infer its existence from its observable effects in the natural order. Besides, as noted again below, mind is something that exists in the natural order, so it is not pure hypothesis. The realities to be explained are givens that cannot be avoided or put on hold under the vacuous claim that it has “not yet” been worked out how such and such can or did happen. That is like saying we have not yet figured out how to get pigs to fly (and still be pigs). Appealing to the “super”-natural must happen, and that appeal must be to adequate “super”-natural cause right now. That point can serve as a kind of “theistic proof,” a proof recognizable from observable effects. Theistic proofs historically have claimed it is more sensible to believe in “god” than not to do so. Philosophers will say that the argument from design and purpose (teleological) as well as necessary, adequate source for beginning (cosmological) are offered strictly as general claims about probability more than necessity. The origin-design-purpose set becomes “apparent” instead of necessary in the minds of evolutionary theorists, because the form might supposedly come by chance from mutation acted on by environment, and such a form might suffice to accomplish a result, that is, an “apparent” result since what looks like a “result” can come about without a purpose and therefore without a god to purpose it.
To typical Darwinian atheists and agnostics, the “theistic proof” implicit in Question #1 and Question #2 leads not to necessity, but probability. Objecting to arguments from irreducible minimums or complexities frequently, then, takes the form of xnth 4 or sheer mystery till future research discovers how it happened or could have happened. Though the proposed number is unimaginably astronomical, to their way of thinking, it represents improbability not actual impossibility. It is imaginable, theoretically, for a person the next day to find on Baker Street, London, the wedding ring she lost overboard in the mid-Pacific. A theorist could postulate as yet unknown technical advances to make that happen—processes for distant detection and retrieval in water and darkness as well as sufficiently speedy travel. Such postulated processes are conceivable because they merely increase in degree or number processes already in existence. So, on that kind of analogy no matter how improbable, the Darwinian evolutionist may prefer to claim that virtually boundless time and space make the improbable even necessary.
This study, however, posits that reciprocationʹ at the level of a whole living entity makes infinite space and time irrelevant. Lengthening the time and broadening the area for straight-line natural causation cannot approach the essential feature in living bodies: reciprocalityʹ, a causal pattern parallel in kind to any form of straight-line causality. With living entities there is observable reciprocalityʹ from any starting point back around the cycle to that point for what is necessary for the entity’s life and functions. Such a system originally cannot come-to-be in sequence by increments because any separate, originating part itself would not be living; it would not be living because, being outside the system, it would not have what it had to be getting from elsewhere in the system for being alive and producing what likewise those other subsequent parts need. Furthermore, it would not be connected to the entity, etc. Real reciprocalityʹ within a living physical entity eliminates external material contribution. If irreducible complexity at the cell level implies intelligence by virtue of sets that must already be in complex combination to produce any effects, more observably so does reciprocal causationʹ in a complex, closed, living loop at the level of the whole. At least, such an arrangement implies inference to the best adequate explanation; but the claim in this presentation is that only intelligent design suffices to adequately account for the real phenomena in the biosphere, particularly reciprocationʹ.
The issue is this: what makes the result occur? Time is not a cause; neither is space. They only make room for a cause.
We know from scientific observation that the organs in a living body can produce what they produce.
We know from scientific observation that what the necessary parts produce, do in fact make possible what are necessary for functional viability all the way around the loop back to itself—as well as crosswise to any “dead ends.”
We know from observation that the biosphere has series, sets, and cycles of causality that include direct and indirect reciprocalityʹ.
We know as well from experience that there is such a thing as mind–however it is explained—and that mind-will-authority can design complex entities of the sort that cannot originate themselves.
It remains only to observe that all the living loops and their parts are necessary from the start (simultaneous origin).
That in turn requires adequate, external cause since internal cause has already been judged inadequate—which is the same idea behind “theistic” proof. The “proof” in this case, however, must be more than apparent, because there is no other possible source internally besides the reciprocatingʹ ones. (The number of internal causes is finite as well as specific within complexity.) Any apparent proof would only be a kind of straight-line causality because it is the only other kind of causality.
So,
this “theistic proof” establishes necessary existence of “god” external to the
biosphere, not just probable existence. It seems more than a stretch to call
merely “improbable” the 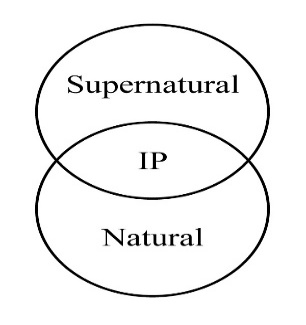 accidental/random/chance
confluence hierarchies of myriads of reciprocatingʹ
parts, compenetrating, and interdependent all the way from the base matter in
cells up through the levels of reciprocatingʹ organs
in complete entities. Time (in minutes) and death (inexorably) preclude
improbability as a relevant description in viable systems. The result must be instantaneous,
external to the physical order, and adequate to produce “from scratch” (no
prehistory) the cell, life’s most basic, unimaginably complex unit, which lies
at the bottom of the “simple”-to-complex system.
accidental/random/chance
confluence hierarchies of myriads of reciprocatingʹ
parts, compenetrating, and interdependent all the way from the base matter in
cells up through the levels of reciprocatingʹ organs
in complete entities. Time (in minutes) and death (inexorably) preclude
improbability as a relevant description in viable systems. The result must be instantaneous,
external to the physical order, and adequate to produce “from scratch” (no
prehistory) the cell, life’s most basic, unimaginably complex unit, which lies
at the bottom of the “simple”-to-complex system.
Whereas there may be some desire to reserve “impossible” for metaphysical processes, it seems appropriate to use “impossible” for something in any system relative to the real features of that system. It is a kind of category error to limit impossible to the metaphysical when speaking about the workings of the physical. What can or cannot happen needs to be understood in terms of the features of the system under consideration. Besides, in the Judeo-Christian worldview, God created mankind in his own image. (See the preceding diagram.) The implication is that God and humankind share significant InterPersonal capacities (“Let us make . . . male and female”; Genesis 1:26-27); hence, natural and supernatural overlap in their features. So possible and “impossible” can apply at least to the shared features in the overlap—as in the case of intelligence and will.
As a result, in the physical system there is as much propriety in saying it is “impossible” (a) for the same person to run and crawl at the same time in the same place, not just as people use the words “run” and “crawl,” but as to the real actions themselves (mutually exclusive). As regards time-space reality, (b) out of the infinite possibilities that are capable of occurring at a given time and place, all the other possibles are eliminated by the one that happens. Furthermore, it is impossible for “pigs to fly.” As regards the built-in features of an entity, (c) out of all the kinds of actions that could happen, all the others are eliminated by the one(s) that entity is designed to do. In that same vein, by the nature of the case, in the biosphere
it is inherently impossible for material nature unaided to simultaneously originate reciprocatingʹ parts that each together produce all the results that are necessary for the ongoing viability of any whole organism and its other parts.
Since the system is closed and reciprocalʹ, there is no place of entry for external, natural cause. By the nature of the case, no prior, external, physical cause can bring about simultaneously a closed loop of necessary reciprocating causesʹ together with all the “embedded” features in the lists under Questions #1 and #2 above. To reiterate, by the nature of reciprocal causationʹ in a closed living loop, it is no more possible for such an entity to originate naturally than it is for pigs to fly. We feel justified in reverting to the age-old intuitive observation, “From nothing nothing comes” (nihil ex nihilo).
The line of thought here serves as a kind of teleological argument, but it is a teleological argument rooted in reciprocal causationʹ rather than in straight-line causal series, as traditional formulations seem to be. The reciprocatingʹ process inside biological nature requires reaching outside of nature to get sufficient cause. That differs from origin-design-purpose from First Cause, which might be conceived of as inside the same realm as its derivatives (non-transcendence), such as could be the case in pantheism, panentheism, or eternal materiality. Reciprocal causationʹ in a closed living loop eliminates that option.
Suffice it to say, this “theistic proof” is like the others in being able to imply only some characteristics of “god”: surely intelligence, volition, transcendence (in kind) to material reality, imminence (as nearness) to the material realm, imperceptibility/spirituality/non-materiality; somewhat sufficient for omni-potence/authority and omni-science to bring about the result (at least omnipresence via omniscience); consciousness and (semi)personhood; perhaps eternality, aseity (self-existence), sovereignty, uniqueness. We are not likely to reason interpersonalism, love, immutability, and monotheism into it, which are requisite features of the Judeo-Christian Yahveh at least. Those and others of God’s characteristics must come by special revelation experienced in time and history. For that reason, this presentation puts quotation marks around “theistic proof” and uses “god” in lower case. The contribution of these thoughts about science is for standing in contrast to materialistic general evolution more than for discovering God. General revelation through nature may help prepare the human mind for understanding and accepting the special revelation that can come in history.
Summary and Integrations
Observational Reality About the Biosphere
Reciprocal causalityʹ inside a closed living loop
Inferences About Bioentities
Simultaneous origin of vital parts, external causation, and intelligent agency for all bioentities within their viability limits, operational boundaries, and completeness requirements
Integrations
Intelligent design researchers have highlighted irreducible complexities at the biochemical level by drawing attention to (1) the complex sets of components that function jointly to produce their effects, notably at the molecular and cellular levels. These sets also have levels of subunits that must operate as fully formed sets. The systems stack up in complex hierarchies from quarks, protons and electrons, atoms, molecules, and cells toward organs and whole entities.
Reciprocal causalityʹ additionally highlights irreducible complexity at the level of the whole by drawing attention to (1) each part/organ as individually producing what the next components themselves need to live/be before those parts in turn can produce conjointly (2) what the whole entity needs for performing its functions. So, RCʹ stresses another level of hierarchy to the complexity. Not only are there complex series and sets but also complex reciprocalitiesʹ atop the others. In any series, any set, or any reciprocatingʹ system, removing one part makes non-functional the whole series, set, or reciprocatingʹ system.
The above analysis of reciprocalityʹ notes that, among all the other kinds and patterns of processes that take place in a complete entity, A produces what B needs to be/live and then to produce what the living B produces distinct from what B received from A. A does not produce B or part of B or combine with B to produce C. (In the case of the heart-lungs epitome, the lungs add oxygen to the blood mixture in contrast to bringing about a new type of product.) B is already capable of being/living and producing—as long as it receives from A the nutrients/materials necessary for existing and then delivering an effect for A as well as for C, D, E, F, etc., on around the loop.
Together the types of patterns combine two kinds and degrees of complexity within the living whole. (1) One uses combinations that conjointly yield effects; (2) the other uses sequences of effects to produce whole-entity functionality. In both cases, removing even one unit from the set or sequence eliminates the functionality of that set or sequence as well as the next level higher that depends on the lower one.
With these observations in place, “external cause” of an intelligent sort has a guaranteed role in the picture of both levels of reality: supernatural and natural intelligence at the detailed biochemical level, at the level of whole entities, and anything in between. Methodological naturalism can have as large an explanatory role as it can fulfill as per real features of the natural order. Put otherwise, mutation plus natural selection can have as large a place as it can actually occupy. If something at first thought to be supernatural turns out to be natural (or perhaps either, depending), then the “god” factor moves back from direct cause to indirect cause instead of falling out of the picture. Even there, the design element still applies because of the limits of variation built into the entity at the DNA level—another application of “viability limit.” As unguided events without intended goals, random mutations plus natural selection do not build on each other toward destinies as in making series or sets or cyclesʹ; otherwise, they would not be random. At some point, the entity ceases to be functional and reproductive; it dies and goes extinct. The whole biosphere must make sense scientifically top to bottom.
Endnotes
1See PART II.
2For starters, see Michael J. Behe, A Mousetrap for Darwin (Discovery Institute Press) 2020, 555pp.
Michael J. Behe, Darwin’s Black Box: The Biochemical Challenge to Evolution (Free Press) 2006, 329 pp.
Michael J. Behe, The Edge of Evolution: The Search for the Limits of Darwinism (Free Press) 2007, 321pp.
Michael J. Behe, Darwin Devolves: The New Science About DNA That Challenges Evolution (HarperCollins) 2020, 342pp.
Michael J. Behe, William A. Demski, and Stephen C. Meyer, Science and Evidence for Design in the Universe (Ignatius Press) 2000, 234pp.
William A. Dembski, Intelligent Design: The Bridge Between Science and Theology (InterVarsity Press), 1999, 312pp.
Stephen C. Meyer, Signature in the Cell: DNA and the Evidence of Intelligent Design (HarperCollins) 2009, 613pp.
3This theistic process could have two variants, one in which “god” created each kind “from scratch” and another where he created the next kind by modifying a previous one in the tree of life. The second scenario appeals to external cause in place of evolution’s postulated transitional forms in getting from one known point in the tree to the next; this second variant is a form of progressive creationism.
An additional note: in the DNA of each entity there is significant built-in variation potential in, say, dogs, cats, cows, frogs, elephants, birds, fish, grasshoppers. They remain within genetically interbreedable boundaries of variation. There is no way, however, even to imagine a step-by-step set of small random changes to get from a grasshopper to an elephant. Much less is there any way to demonstrate a pathway from one to the other. Imagination is not science; silence is not proof.
To help avoid confusion, we have become accustomed to using “progressive creation” to indicate external cause by God taking place over time. That leaves “theistic evolution” to designate God’s creating into base matter the internal capacity to evolve into all the forms that have arisen since the beginning. It is the kind of format implicit in the expression “God used evolution to create.” In common parlance, however, people may use such language to talk about another arrangement where, say, the car evolved from a Model A to a Cadillac. The cause for the changes lies outside the realm of the cars and in the designers. It is phenomenological language to speak of the car as evolving. The expression “theistic evolution” could be used similarly, of course. The main thing is to keep the realities distinguished in speaking about the process that describes the origin of the biosphere.
4Simple to complex is not exactly “simple.” The complexity level in monocellular life forms and even below is mind-boggling. Michael Behe enumerates nearly twenty precise steps in just getting the vision experience in the brain from a photon hitting the retina in the eye (“Design at the Foundation of Life” in Science and Evidence for Design in the Universe, pp117-18). At the organic molecular level, that is just one aspect of one feature variously designed among a plethora of other constructions for sightedness in organisms. Moving through this simple-to-complex arrangement gets further complicated by needing to cross the inorganic-to-organic threshold, to make the fission-to-fusion switch in reproduction, to assemble coherently the vast number of subunits in a viable entity, and to accommodate multilinear descent features as diverse as in mosaics (see earlier)—and to do it en masse in progress without breaching the viability limit. We may wax bold by saying that God himself could not lay out a pathway from bottom to top in the biosphere if he were to limit himself to the features of the physical order.
Darwin’s idea of evolution by mutation originated at a time (1865) when scientists still believed in the spontaneous generation of life. The mind-blowing complexity of just getting across the inorganic-to-organic threshold has since come to light; yet theorists still hang onto the original simplistic understanding that underlay proposing biological evolution. At least from a distance, it suggests avoidance behavior. Simple, crudely chipped stones found in a dig site in Kenya can be considered tools (design); alongside them hangs a detailed, analyzed picture of the eye attributed to random evolution (chance). Something seems to be at work here besides science.
5Michael J. Behe, Darwin’s Black Box: The Biochemical Challenge to Evolution (Free Press) 2006. Other writers on intelligent design who address this issue include William A. Dembski and Stephen C. Meyer. For an extensive listing of relevant literature prior to 2009, consult Stephen Meyer’s Signature in the Cell: DNA and the Evidence for Intelligent Design (HarperCollins Publishers) 2009, pp, 563-95; also see the endnotes in Michael J. Behe’s A Mousetrap for Darwin (Discovery Institute, Seattle) 2020, pp, 513-46.
The complexity level in straight-line causality in the direction of smallness is evidently not unlimited (Behe, Darvin’s Black Box, p. 13), which is what we would expect in space-time reality. As to time, “the mind rejects an infinite regress,” so also as to size, the mind rejects an infinite smallness. In some way, the regress must end at a “starter”; finiteness must end at a base level that is adequate for explaining everything subsequent. Whether (1) starting a reciprocal causalityʹ set in a closed, living loop, or (2) going backward in a cause-effect straight-line series, or (3) regressing into smallness, physical causation must yield to non-physical mind/choice/authority/action. That is because nothing more basic exists in the physical world to get the series going, or to originate a set functioning only as a set, or to get the reciprocal loop to cycle. Thus, size joins sequence and sets and reciprocalʹ loops as aspects of the cosmological proof.
SUPPLEMENT
This addendum surveys the body of the presentation above to highlight features that in some cases have broader application than the biosphere. Additional considerations come into play that flesh out items in the unified field of knowledge thoughtful persons may want to work on.
There is overlap between natural and supernatural, an overlap consisting of interpersonal features, particularly mind/will/authority/act, also termed “spiritual” features. Inorganic and organic both involve chemical machinery. Fission and fusion reproduction and plant and animal life operate with some of the same biochemistry. Animals and humankind have crossovers in their physical components—and somewhat beyond as manifested in different degrees. The uniqueness of supernatural, human, animal, organic, and inorganic share a certain degree of the less complex level nearest them as the whole system form a kind of “chain of being.”
Reciprocal causalityʹ is the most irreducible form of combinationalism because (a) it is the least capable of receiving external contribution in completing the combination. Furthermore, (b) its time factor requires the most urgent need for all parts in the living entity to be present from start to finish to avoid dissolution—death by virtue of the viability limit.
As a term, “viability limit” has the advantage of forcing time into the process. It stresses the fact that viability cannot wait indefinitely in suspended animation till the vital factor arrives from somewhere else to finish the reciprocatingʹ set—even if some external source existed for it along with some way to import it coherently in the deficient cycle. “Irreducible complexity” has the advantage of describing a wider range of entities than organic ones, and paves the way for appreciating its application in the realm of living things. Combinations, series, sets, and cycles highlight the need for all parts in bringing about the necessary result: life, and it is in biology that evolution is most important and most controversial for understanding our human nature and human experience.
Finally, there is additional food for inquiry. One such matter relates to mutation in larger, multicellular animals that reproduce bisexually by fusion. Research on intelligent design and reports on abiogenesis deal often with monocellular life, which involves asexual reproduction by fission (cell division). How mutational alterations to the DNA get passed on intergenerationally seems relatively straightforward; the whole cell divides to become another example (replication) that carries within it all its previous features as well as the mutational modification.
In multicellular entities, how does mutation operate? Where does it occur? Does it occur in one cell and then spread to all the others? How would it spread? Do not all the parts of the entity carry the DNA distinctive to each entity whether in the skin, the tongue, the brain, blood, or whatever? Does mutation occur in all the cells simultaneously? At any rate, how do mutations in cells get into the reproductive elements in the semen or ovum to pass on to the next generation? Or does mutation always occur directly on the reproductive materials in the male or female? If a mutation occurs in the male or female, how does it match up with the reproductive form (coherence) in the other sex so the combination can “work”? Does the same compatible mutation have to occur in both sexes at the same time and place?
christir.org